研究揭示膜蛋白在水稻乙烯信号转导中的双重作用
中国科学院遗传与发育生物学研究所张劲松教授团队研究揭示了膜蛋白MHZ3协同乙烯受体调控OsCTR2(CONSTITUTIVE TRIPLE RESPONSE2)磷酸化,进而控制水稻乙烯信号转换的机制。
植物无法像动物一样通过移动来逃避不利的环境。相反,它们通过动态调节体内激素水平来适应不断变化的环境。
乙烯是植物中存在的一种典型的气态激素,可在细胞、组织和个体之间自由扩散。短时间的乙烯爆发可能增强植物的适应性,而长时间的乙烯暴露则可能对植物的生长发育有害。乙烯的这些特性表明乙烯信号的启动和终止应该是一个快速的过程。
然而,对于乙烯信号早期事件的理解仍然有限。
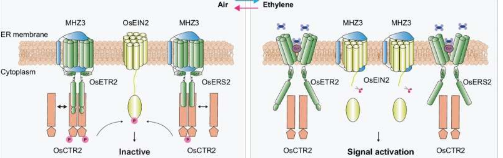
张教授团队利用水稻乙烯信号负调节因子 OsCTR2 在乙烯刺激下快速发生磷酸化转变这一现象,将其作为信号激活的灵敏读数。对乙烯不敏感的水稻突变体 mhz3 的分析表明,MHZ3 与乙烯受体相互作用,促进OsCTR2 的磷酸化。
上位性分析表明MHZ3和乙烯受体在促进OsCTR2磷酸化方面相互依赖。MHZ3和乙烯受体共同定位在内质网膜上。
在空气中,MHZ3与亚家族I和亚家族II乙烯受体相互作用,稳定乙烯受体与OsCTR2的结合,促进OsCTR2附着到内质网膜上,从而维持OsCTR2磷酸化活性并关闭乙烯信号。
乙烯处理会破坏 MHZ3-受体-OsCTR2 复合物内的结合,降低 OsCTR2 磷酸化,并启动下游乙烯信号传导。
该团队前期研究发现,MHZ3是水稻乙烯信号通路中正调控因子OsEIN2的稳定因子,发挥正向调控作用。本次研究进一步发现,MHZ3通过与乙烯受体相互作用,调控OsCTR2的磷酸化,从而参与乙烯信号传导的早期事件,发挥负向调控作用。
研究阐明了MHZ3在乙烯信号通路中的双重作用,在缺乏乙烯的情况下,MHZ3与乙烯受体相互作用,维持OsCTR2磷酸化,使OsCTR2保持活性,乙烯信号关闭。
在乙烯存在的情况下,MHZ3与受体之间的相互作用减弱,导致OsCTR2失活,而MHZ3则重新定向并稳定OsEIN2,使乙烯信号保持活跃。
这项研究为乙烯信号级联的早期事件提供了重要见解,也可能提供适用于其他信号通路的概念范例。
免责声明:本文由用户上传,与本网站立场无关。财经信息仅供读者参考,并不构成投资建议。投资者据此操作,风险自担。 如有侵权请联系删除!
-
【木地板材料】木地板是一种广泛应用于家居和商业空间的地面装饰材料,因其自然美观、耐用性强、环保性好等特...浏览全文>>
-
【木地板安装】木地板安装是一项需要细致规划和专业操作的工程,无论是新房装修还是旧房翻新,选择合适的木地...浏览全文>>
-
【木的组词怎么写】在汉语学习中,词语的积累和运用是非常重要的部分。对于“木”这个字来说,它不仅是常见的...浏览全文>>
-
【木的象形字有哪些字】在汉字的发展过程中,许多字最初都是通过象形的方式创造出来的,用来表示具体的事物。...浏览全文>>
-
【木岛法子介绍】木岛法子(Kazuko Kikuchi)是日本著名演员、模特及艺人,以其在影视作品中的出色表现和独特...浏览全文>>
-
【木代尔是什么面料】“木代尔是什么面料”是许多消费者在选购衣物时常常会提出的问题。木代尔是一种天然纤维...浏览全文>>
-
【木代尔和莫代尔哪种面料好】在选择衣物面料时,很多人会遇到“木代尔”和“莫代尔”这两个名称,容易混淆它...浏览全文>>
-
【萝卜的营养价值】萝卜是一种常见的根茎类蔬菜,不仅味道清脆爽口,还具有丰富的营养价值。无论是生吃、炒食...浏览全文>>
-
【萝卜的药用功效和作用】萝卜,作为日常生活中常见的蔬菜之一,不仅味道清脆、营养丰富,还具有多种药用价值...浏览全文>>
-
【萝卜的家常做法】萝卜是一种非常常见的蔬菜,不仅价格实惠,而且营养丰富,适合多种烹饪方式。无论是炖、炒...浏览全文>>